




Las unidades fotosintéticas de las plantas verdes: los cloroplastos (continuación)
Características estructurales de los cloroplastos (continuación)
Los cloroplastos de la mayoría de las plantas superiores tienen unas regiones, llamadas «granas», en el que los tilacoides están firmemente apilados. Cuando se observan al microscopio electrónico en un ángulo oblicuo, las granas se muestran como discos apilados, y cuando se observan en sección transversal, se evidencia que algunos tilacoides se extienden desde una grana a otra a través del estroma.
Se estima que los delgados espacios acuosos dentro de los tilacoides permiten mantener conectados entre sí los tilacoides dentro del estroma. Estos espacios de los tilacoides están aislados de los espacios del estroma mediante laminillas relativamente impermeables.
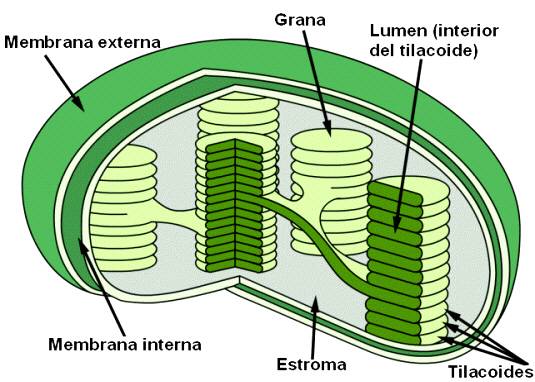
Estructura de un cloroplasto
Las reacciones de la luz se producen exclusivamente en los tilacoides. Se requiere la compleja organización estructural de las láminas para que su función sea adecuada. Los tilacoides que estén aparentemente intactos son necesarios para la formación de ATP, pero los que se han disgregado en unidades más pequeñas ya no pueden formar ATP, incluso aunque la conversión de luz en energía química se produzca durante el transporte de electrones en estas unidades. Tales fragmentos laminares pueden llevar a cabo la reacción de Hill, con la transferencia de electrones desde el agua al NADP+ (La nicotinamida adenina dinucleótido fosfato, en su forma reducida).
Composición química de las laminillas de los cloroplastos
Lípidos laminares
Las laminillas consisten en cantidades aproximadamente iguales de lípidos y proteínas. Alrededor de un cuarto de la porción lipídica de las laminillas se compone de pigmentos y coenzimas; el resto son diversos lípidos, incluyendo los compuestos polares, tales como fosfolípidos y galactolípidos.
Estas moléculas de lípidos polares tienen grupos de «cabezas» que atraen el agua (es decir, son hidrófilos) y ácidos grasos «colas» que son solubles en aceite y repelen el agua (es decir, son hidrófobos). Una segunda capa de fosfolípidos establecen una bicapa en donde las cabezas hidrófilas están en contacto con la solución acuosa a cada lado de la bicapa.
Intercalado entre las cabezas están las colas hidrofóbicas, creando un ambiente hidrofóbico donde el agua queda excluida. Esta bicapa lipídica es una característica esencial de todas las membranas biológicas.
Las membranas laminares pueden funcionar como material eléctrico aislante y permitir una carga, o diferencia de potencial, para desarrollarse a través de la membrana. Una carga de este tipo puede ser una fuente de energía química o eléctrica.
Aproximadamente una quinta parte de los lípidos laminares son moléculas de clorofila; el tipo de clorofila «a» es más abundante que el segundo tipo, la clorofila «b». Las moléculas de clorofila se unen específicamente a pequeñas moléculas de proteínas. La mayoría de estas proteínas de clorofila son pigmentos «captadores de luz». Éstos absorben la luz y pasan su energía a la clorofila, a moléculas especiales que están directamente involucradas en la conversión de la energía luminosa en energía química. Cuando una de estas moléculas especiales de la clorofila es excitada por la energía de la luz, cede un electrón.
Hay dos tipos especiales de moléculas de clorofila: Una, la llamada «P680», tiene un pico de absorción en el espectro de 684 nanómetros; la otra, llamada «P700», muestra un pico de absorción en el espectro de 700 nanómetros.
Aunque las clorofilas son las principales moléculas que absorben la luz en las plantas verdes, otros pigmentos tales como carotenos y carotenoides también pueden absorber la luz, y pueden complementar la clorofila como moléculas que absorben luz en algunas células de la planta. La energía de la luz absorbida por estos pigmentos debe pasar a la clorofila antes de que ocurra la conversión en energía química.
Proteínas laminares
Muchas de las proteínas laminares son componentes de los complejos proteínicos de la clorofila ya descritos antes. Otras incluyen enzimas y coenzimas que contienen proteínas. Las enzimas se requieren como catalizadores para reacciones orgánicas específicas dentro de las laminillas. Las coenzimas de proteínas, también llamadas cofactores, incluyen importantes moléculas portadoras de electrones llamados «citocromos»; son proteínas que desempeñan un papel vital en el transporte de la energía química.
En las proteínas con un peso molecular total de 900.000, hay dos átomos de manganeso, 10 átomos de hierro, y seis átomos de cobre. Se requieren estos átomos de metal para la actividad catalítica de algunas de las enzimas importantes en la fotosíntesis. Los átomos de manganeso están involucrados en la disociación del agua y la formación de oxígeno. Tanto el cobre como las proteínas que contienen hierro funcionan en el transporte de electrones en la etapa final de la fotosíntesis; una de las proteínas que contiene hierro es la llamada ferredoxina; es un componente soluble en los cloroplastos.
Una de las proteínas que contiene cobre es la llamada plastocianina (PC). Son moléculas solubles en agua y pueden moverse a través del espacio interior de los tilacoides, llevando electrones de un lugar a otro.
Quinonas laminares
Las moléculas pequeñas llamadas plastoquinonas se encuentran en número sustanciales en las laminillas. Al igual que los citocromos, las quinonas tienen papeles importantes en el movimiento de electrones entre los componentes de las reacciones luminosas. Puesto que son solubles en lípidos, pueden difundirse a través de la membrana. Ellos pueden portar uno o dos electrones y, en su forma reducida (con electrones añadidos), llevan átomos de hidrógeno que pueden ser liberados como iones de hidrógeno cuando los electrones agregados se transmiten, por ejemplo, a un citocromo.
